 W
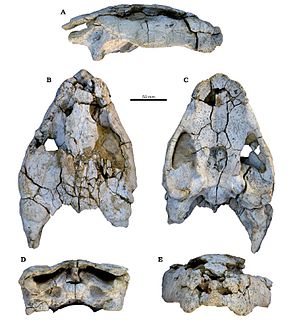
WAegisuchus – rodzaj krokodylomorfa z rodziny Aegyptosuchidae żyjącego w późnej kredzie w północnej Afryce. Został opisany w 2012 roku przez Caseya Hollidaya i Nicholasa Gardnera na podstawie puszki mózgowej dużego osobnika. Nazwa rodzajowa pochodzi od greckich słów aegis („tarcza”) i souchos („krokodyl”), co odnosi się do ornamentacji czaszki Aegisuchus. Epitet gatunkowy gatunku typowego honoruje Lawrence'a Witmera, którego badania i pomoc pozwoliły zidentyfikować okaz Aegisuchus.
 W
WAlanqa – rodzaj pterozaura z rodziny azdarchidów (Azhdarchidae) żyjącego na początku późnej kredy na obecnych terenach Afryki. Został opisany w 2010 roku przez Nizara Ibrahima i współpracowników w oparciu o pięć fragmentów szczęk odkrytych w złożach Kem Kem w Maroku. Holotypem jest spojenie żuchwy oznaczone numerem katalogowym FSAC-KK 26. Dwa fragmenty zostały po raz pierwszy opisane w 1999 roku przez Petera Wellnhofera i Érica Buffetauta i wstępnie zidentyfikowane jako należące do pteranodontów, jednak z taką interpretacją nie zgodzili się Aleksander Awerianow i współpracownicy, którzy zasugerowali, że skamieniałości te należą do azdarchidów. W Kem Kem odnaleziono również szczątki szkieletu pozaczaszkowego azdarchidów – Ibrahim i in. nie zdecydowali się przypisać ich do Alanqa saharica, jednak ich zdaniem hipoteza taka jest prawdopodobna. Holotyp składa się z trzech sąsiadujących fragmentów o łącznej długości 344 mm – długość brakującej części oszacowano na podstawie porównań z innymi azdarchidami na około 80 mm. Długość żuchwy FSAC-KK 26 wynosiła prawdopodobnie około 67,5 cm. Większość szczątków odnalezionych w Kem Kem – w tym również holotyp – należała do osobników o rozpiętości skrzydeł wynoszącej 3–4 m, jednak fragment dużego kręgu szyjnego, prawdopodobnie również należący do Alanqa saharica, sugeruje, że przedstawiciele tego gatunku mogli osiągać nawet 6 m rozpiętości skrzydeł. Szczęki były proste i ostro zakończone, podobnie jak należące do rodzajów Zhejiangopterus i Quetzalcoatlus, co wspiera hipotezę o przynależności A. saharica do Azhdarchidae. Nie odnaleziono żadnych zębodołów, co dowodzi, że Alanqa była bezzębna. Szczęki prawdopodobnie pokrywała ramfoteka.
 W
WAnatosuchus – rodzaj niewielkiego krokodylomorfa z grupy Mesoeucrocodylia żyjącego we wczesnej kredzie na terenie dzisiejszej Afryki. Prawdopodobnie był drapieżnikiem poruszającym się na wyprostowanych kończynach i polującym na bezkręgowce i niewielkie wodne kręgowce. Pozycja filogenetyczna rodzaju jest niepewna – analizy wskazują na jego przynależność do Notosuchia lub Neosuchia.
 W
WAngolachelys – rodzaj żółwia skrytoszyjnego żyjącego w późnej kredzie na obecnych terenach Afryki. Został opisany przez Octávio Mateusa i współpracowników w 2009 roku w oparciu o niemal kompletną czaszkę, paliczek, dwa kręgi szyjne, proksymalną część kości biodrowej, fragmenty pancerza oraz niezidentyfikowane elementy. Skamieniałości odkryto w pobliżu miejscowości Iembe w angolskiej prowincji Bengo. Pochodzą one ze złóż Tadi datowanych na górny turon, około 90 mln lat. Miejsce, w którym odnaleziono szczątki, znajduje się w pobliżu lokalizacji typowej mozazaura Angolasaurus bocagei. Czaszka Angolachelys mbaxi jest dłuższa niż szersza i stosunkowo duża – od kłykcia potylicznego od czubka pyska mierzy 179 mm, a szerokość na wysokości kości kwadratowych wynosi 137 mm. Według analizy filogenetycznej przeprowadzonej przez autorów Angolachelys jest przedstawicielem grupy Eucryptodira należącym do kladu Angolachelonia – jego taksonem siostrzanym jest żyjąca w późnym apcie na obecnych terenach Wielkiej Brytanii Sandownia. Angolachelys reprezentuje jedną z pierwszych linii ewolucyjnych owodniowców, której przedstawiciele zasiedlili południowy Atlantyk po oddzieleniu się Afryki od Ameryki Południowej.
 W
WArambourgisuchus – rodzaj krokodylomorfa z rodziny Dyrosauridae. Żył w późnym paleocenie na terenie dzisiejszego państwa Maroko. Odnaleziono go dzięki współpracy Francji i Maroka w roku 2000 w regionie Sidi Chenane. Zwierzę opisał 5 lat później paleontolog Stéphane Jouve.
 W
WAsilisaurus – rodzaj dinozauromorfa z grupy Silesauridae żyjącego w środkowym triasie na obecnych terenach Afryki. Został opisany w 2010 roku przez Sterlinga Nesbitta i współpracowników w oparciu o przednią część kości zębowej wraz z zębami odkrytą w datowanych na anizyk osadach ogniwa Lifua w złożach Manda w pobliżu tanzańskiego miasta Litumba Ndyosi. Odnaleziono jednak izolowane szczątki należące do co najmniej czternastu osobników, składające się w sumie na niemal kompletny szkielet – jedynie niektóre części czaszki oraz nadgarstki pozostają nieznane. Według Nesbitta i współpracowników (2010) większość skamieniałości należała do osobników młodocianych, które osiągnęły 60–80% maksymalnych rozmiarów; na tej podstawie, oraz na podstawie porównań ze szkieletem silezaura, autorzy szacowali, że dorosli przedstawiciele gatunku A. kongwe osiągali około 2–3 m długości ciała. Czaszka ma podobne proporcje jak czaszka silezaura i podobnie jak u niego oraz u Sacisaurus kość zębowa jest ostro zakończona. Szacowana długość kości ramiennej i promieniowej sugeruje, że Asilisaurus miał wydłużone kończyny przednie, jak Silesaurus. Panewka stawu biodrowego była zamknięta, jak u wszystkich przedstawicieli Ornithodira nienależących do dinozaurów. Kość skokowa i kość piętowa to u Asilisaurus odrębne struktury, w przeciwieństwie do pterozaurów, teropodów i Lagerpetidae, u których były one na wpół skostniałe.
 W
WAustralochelys – rodzaj wymarłego żółwia, najstarszego jakiego znaleziono w Afryce, żyjącego na przełomie późnego triasu i wczesnej jury.OpisZnaleziono tylko czaszkę oraz fragment pancerza, które opisano pod nazwą Australochelys africanus. Trudno ocenić wiek znaleziska, dlatego często przyjmuje się, że jest to wczesna jura. Skamieniałości gatunku A. africanus porównuje się do znalezionych w Ameryce Południowej Palaeochersis talampayensis. Pomimo tak skąpych znalezisk odkrycie tych skamieniałości jest bardzo ważne, gdyż jak oceniają naukowcy w tym okresie zaczął się podział na żółwie bokoszyjne (Pleurodira) i skrytoszyjne (Cryptodira).Miejsce znalezieniaprowincja Wolne Państwo, RPA (Afryka)
 W
WAzendozaur (Azendohsaurus) – rodzaj roślinożernego archozauromorfa żyjącego w późnym triasie na obecnych terenach Afryki. Gatunek typowy rodzaju, Azendohsaurus laaroussi, został opisany na podstawie kości zębowej zawierającej zęby oraz izolowanych zębów. Później przypisano do niego także inne znalezione w Maroku szczątki, złożone z kręgów piersiowych, kości obręczy barkowej oraz przednich i tylnych kończyn. Początkowo na podstawie podobieństw do zębów fabrozaura i likorina uznany za roślinożernego dinozaura ptasiomiednicznego, jednak późniejsi autorzy sugerowali, że jest on prozauropodem. Nowsze badania wskazują jednak, że Azendohsaurus nie należy do dinozaurów ani w ogóle archozaurów, lecz jest bazalnym archozauromorfem, być może blisko spokrewnionym z trilofozaurami. Stosunkowo kompletne szczątki pochodzące z przełomu środkowego i późnego triasu Madagaskaru, opisane w 2010 roku jako Azendohsaurus madagaskarensis, potwierdzają status azendozaura jako bazalnego archozauromorfa, u którego występowały niektóre cechy uznawane wcześniej za diagnostyczne dla zauropodomorfów. Szczątki A. laaroussi pochodzą z datowanych na karnik osadów formacji Argana w górach Atlas, w pobliżu wioski Azendoh w regionie Marrakesz-Tansift-Al-Hauz. Z tej samej formacji pochodzą odciski stóp przypisywane ptasiomiednicznym i teropodom. Azendozaur osiągał prawdopodobnie około 1,8 m długości.
 W
WChenanisuchus – rodzaj krokodylomorfa z rodziny Dyrosauridae. Żył od późnej kredy do późnego paleocenu w północnej Afryce. Jego pozostałości znaleziono w okolicy Sidi Chenane w Maroku. Odkryto je w 2000, a opisano w 2005 roku.
 W
WCrocodylus anthropophagus – gatunek wymarłego krokodyla żyjącego na przełomie pliocenu i plejstocenu we wschodniej Afryce. Został opisany w 2010 roku przez Christophera Brochu i współpracowników w oparciu o niekompletną czaszkę oraz fragmentaryczne pozostałości szkieletu pozaczaszkowego (NNHM-OLD-1001) odkryte w 2007 roku w Wąwozie Olduvai, na powierzchni pomiędzy dwiema warstwami tufu wulkanicznego. Oprócz holotypu odnaleziono także szczątki co najmniej kilku osobników, obejmujące pozostałości czaszki, w tym puszki mózgowej, kości kończyn, kręgi, osteodermy. Skamieniałości liczą około 1,84 mln lat – w tym okresie na terenie Wąwozu Olduvai żyły również hominidy Homo habilis i Paranthropus boisei. Crocodylus anthropophagus cechował się dużymi wyrostkami na kościach łuskowych. Nie występowało u niego wydłużenie grzebienia przedoczodołowego typowe dla indopacyficznych przedstawicieli rodzaju Crocodylus oraz środkowych guzków rostralnych, diagnostycznych dla neotropikalnych Crocodylus. Budowa szkieletu pozaczaszkowego jest podobna jak u współczesnych przedstawicieli tego rodzaju. Głęboki pysk i rogi upodabniają C. anthropophagus do Voay robustus – dużego krokodyla żyjącego na Madagaskarze jeszcze około 2000 lat temu. C. anthropophagus osiągał prawdopodobnie rozmiary zbliżone do krokodyla nilowego i był największym drapieżnikiem występującym na terenie Wąwozu Olduvai. Ślady zębów odnalezione na kościach dowodzą, że polował m.in. na wczesne hominidy. Ponieważ współczesne mu hominidy były stosunkowo niewielkie, ślady ukąszeń na kościach prawdopodobnie są wynikiem ataku krokodyli niewielkich lub średnich rozmiarów, gdyż większe osobniki były w stanie zjeść hominida w całości, nie pozostawiając żadnych śladów. Do faktu zjadania człowiekowatych odnosi się nazwa gatunkowa anthropophagus, pochodząca od greckich słów anthropos („człowiek”) i phagus („pożeracz”).
 W
WCrocodylus thorbjarnarsoni – gatunek wymarłego gada z rodziny krokodylowatych (Crocodylidae) żyjący przez większość pliocenu i we wczesnym plejstocenie we wschodniej Afryce.
 W
WDiodorus – rodzaj archozaura z grupy Dinosauriformes żyjącego w późnym triasie na obecnych terenach Maroka.
 W
WDyrosaurus – rodzaj eoceńskiego krokodylomorfa zamieszkującego tereny Afrykę Północną.
 W
WElosuchus – wymarły rodzaj krokodylomorfa. Zamieszkiwał północną Afrykę w epoce wczesnej kredy. E. cherifiensis występował w dwóch pierwszych z wymienionych państw. Pierwotnie włączono go do innego rodzaju – Thoracosaurus (Lavocat). E. felixi żył na terenie dzisiejszego Nigru. Nadmienić należy, że Elosuchus został oddzielony od wspomnianego już drugiego rodzaju przez Broina w 2002. Człowiek ten stworzył nawet oddzielną rodzinę Elosuchidae. obejmującą także również pochodzącego z Nigru krokodylomorfa o nazwie Stolokrosuchus.
 W
WGigantophis garstini – gatunek prehistorycznego węża zaliczany do rodziny Madtsoiidae, jedyny przedstawiciel rodzaju Gigantophis; być może jeden z największych gatunków węży w historii. Prawdopodobnie, jak współczesne węże dusiciele, posiadał w pobliżu kloaki parę niewielkich „ostróg”, przekształconych kości udowych połączonych z resztkami bioder, będących pozostałościami kończyn tylnych. Gigantophis żył 40-38 milionów lat temu w eocenie na terytorium dzisiejszego Egiptu i Algierii.
 W
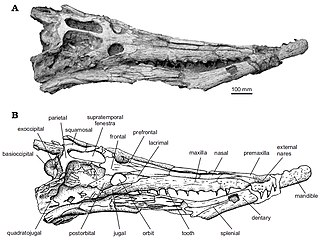
WHamadasuchus – rodzaj krokodylomorfa z grupy Peirosauridae żyjącego w późnej kredzie na terenach współczesnej Afryki. Gatunek typowy rodzaju, Hamadasuchus rebouli, został opisany w 1994 roku przez Érica Buffetauta w oparciu o niekompletną lewą kość zębową z sześcioma zębami odkrytą w datowanych na alb–cenoman osadach na pustyni Hamada w południowym Maroku. W 1999 roku Larsson i Sidor opisali zęby H. rebouli pochodzące z górnych warstw Kem Kem w Maroku. Sugerują one, że Hamadasuchus miał stosunkowo heterodontyczne uzębienie, z trzema występującymi u niego morfotypami. W 2007 roku Larsson i Sues opisali znakomicie zachowane skamieniałości kilku okazów, w tym niemal kompletną czaszkę dużego osobnika, co pozwoliło na dokładniejsze poznanie osteologii czaszki Hamadasuchus i jego pozycji filogenetycznej. Widziana od strony grzbietowej czaszka przypomina trójkąt o wydłużonych ramionach. Cechuje się ona długim pyskiem – u ROM 52620 stanowił około 70% długości czaszki, mierząc 22,8 cm z około 32,5 cm. Na lewej stronie czaszki ROM 52620 nie występuje okno przedoczodołowe, jednak niewielki, mierzący około 6 mm średnicy otwór znajduje się na jej prawej stronie.
 W
WHyposaurus – rodzaj krokodylomorfa z rodziny Dyrosauridae. Gatunek typowy, H. rogersii, został opisany w 1849 roku przez Richarda Owena w oparciu o skamieniałości pochodzące z górnej kredy Ameryki Północnej. Drugim gatunkiem z tego kontynentu jest H. natator, nazwany w 1925 roku przez Troxella na podstawie szczątków odkrytych w New Jersey i datowanych na mastrycht. W 1886 roku Edward Drinker Cope opisał gatunek H. derbianus w oparciu o skamieniałości odnalezione w Brazylii. Cope twierdził, że pochodzą one z kredowych osadów stanu Pernambuco, jednak nie podał dokładnej lokalizacji, wobec czego możliwe, iż zostały wydobyte z osadów formacji Maria Farinha, którą obecnie datuje się na paleocen. W 1930 roku Swinton opisał dwa paleoceńskie gatunki z terenów Nigerii – H. nopcsai i H. wilsoni, klasyfikując je w rodzajach, odpowiednio, Sokotosaurus i Wurnosaurus. W 2007 roku oba zostały uznane przez Stéphane'a Jouve'a za nomina dubia, niemożliwe do odróżnienia od siebie, a także od form północnoamerykańskich. Hyposaurus należy do rodziny Dyrosauridae – analizy filogenetyczne sugerują, że jest stosunkowo zaawansowanym przedstawicielem tego kladu.
 W
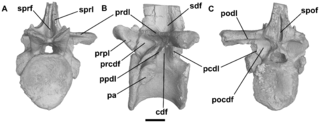
WHypselorhachis – rodzaj rauizucha należący do rodziny Ctenosauriscidae. Żył w środkowym triasie na obecnych terenach Afryki. Jego skamieniałości odkryto w datowanych prawdopodobnie na późny anizyk osadach ogniwa Lifua w złożach Manda w Tanzanii. Holotypem jest jeden kręg grzbietowy charakteryzujący się bardzo wydłużonym wyrostkiem kolczystym, co najmniej 5,5 raza dłuższym od trzonu kręgu. Jedyna autapomorfia gatunku typowego rodzaju, Hypselorhachis mirabilis, jest związana z morfologią wyrostka stawowego przedniego. Hypselorhachis przypominał inne środkowotriasowe archozaury z wydłużonymi wyrostkami kolczystymi, takie jak arizonazaur, lotozaur i Ctenosauriscus, dlatego Butler i in. (2009) przypuszczali, że taksony te mogą tworzyć klad Ctenosauriscidae, jednak nie byli tego pewni. Późniejsze analizy kladystyczne przeprowadzone przez Butlera i wsp. (2011) potwierdziły monofiletyzm Ctenosauriscidae i przynależność Hypselorhachis do tego kladu.
 W
WKaprosuchus – rodzaj krokodylomorfa z rodziny Mahajangasuchidae żyjącego w późnej kredzie na obecnych terenach Afryki. Został opisany w 2009 roku przez Paula Sereno i Hansa Larssona w oparciu o niemal kompletną czaszkę pochodzącą z osadów formacji Echkar w Nigrze datowanych na cenoman, około 95 mln lat. W pobliżu odnaleziono szczątki innego przedstawiciela Crocodyliformes z rodzaju Laganosuchus, teropodów rugopsa, spinozaura, karcharodontozaura oraz zauropodów z grupy tytanozaurów i nienazwanego przedstawiciela rebbachizaurów. Długość czaszki od kości przedszczękowej do kłykcia kości kwadratowej wynosi 507 mm, zaś długość żuchwy – 603 mm. Całe zwierzę osiągało prawdopodobnie około 6 m długości. Kaprosuchus miał ostre, powiększone zęby przypominające kły, osadzone parami w silnie umięśnionych szczękach. Prawdopodobnie mógł szybko i szeroko rozwierać szczęki. Różnice w wielkości poszczególnych zębów są nietypowe dla Crocodyliformes. Złączone kości nosowe sugerują, że przód pyska był wzmocniony, by lepiej wytrzymywać naciski wywoływane silnymi ukąszeniami, oraz zgrubiały i pokryty bruzdami, za życia zwierzęcia przypuszczalnie pokryty rogową pochwą. Oczodoły były skierowane bardziej na boki niż ku górze, co wskazuje, że dorosłe Kaprosuchus nie polowały w wodzie. Sereno i Larsson sugerują, że Kaprosuchus był głównie lub wyłącznie lądowym drapieżnikiem. Prawdopodobnie polował na duże lądowe kręgowce, takie jak dinozaury. Według analizy filogenetycznej przeprowadzonej przez Sereno i Larssona najbliższym krewnym Kaprosuchus jest Mahajangasuchus, wraz z którym należy do kladu Mahajangasuchidae.
 W
WLaganosuchus – rodzaj krokodylomorfa z rodziny Stomatosuchidae żyjącego w późnej kredzie na obecnych terenach Afryki. Został nazwany w 2009 roku przez Paula Sereno i Hansa Larssona. Gatunkiem typowym jest Laganosuchus thaumastos, opisany w oparciu o niemal kompletną żuchwę pochodzącą z osadów formacji Echkar w Nigrze datowanych na cenoman, około 95 mln lat. W pobliżu odnaleziono szczątki innego przedstawiciela Crocodyliformes z rodzaju Kaprosuchus, teropodów rugopsa, spinozaura, karcharodontozaura oraz zauropodów z grupy tytanozaurów i nienazwanego przedstawiciela rebbachizaurów. Żuchwa L. thaumastos mierzy 84 cm długości, co sugeruje, że należała do zwierzęcia o długości 4–6 metrów. Drugi gatunek należący do rodzaju Laganosuchus, L. maghrebensis, został opisany w oparciu o niekompletną lewą kość zębową z czterema zębodołami pochodzącą z osadów złóż Kem Kem w Maroku, również datowanych na około 95 mln lat. L. maghrebensis osiągał mniejsze rozmiary niż L. thaumastos, był jednak do niego bardzo podobny. Niewielkie różnice w budowie kości zębowych potwierdzają jednak ich odmienność na poziomie gatunkowym. Laganosuchus miał bardzo płaskie, wydłużone szczęki w kształcie litery U. Prawdopodobnie nie był w stanie gwałtownie ich otwierać ani zamykać. Sereno i Larsson sugerują, że Laganosuchus i jego najbliższy krewny, Stomatosuchus, były aktywnymi drapieżnikami polującymi z zasadzki na wodne kręgowce. Wiele cech zaawansowanych, takich jak znaczące wydłużenie szczęk, sugeruje bliskie pokrewieństwo Laganosuchus i Stomatosuchus. Według analizy filogenetycznej przeprowadzonej przez Sereno i Larssona Laganosuchus jest najbliższą grupą zewnętrzną kladu obejmującego Bernissartia, Isisfordia oraz krokodyle.
 W
WMezozaur – rodzaj żyjącego w karbonie i permie, około 300 mln lat temu bazalnego zauropsyda zaliczanego do anapsydów. Jedyny znany przedstawiciel rzędu Mesosauria.
 W
WMilleretta (Milleretta) - rodzaj niewielkiego zauropsyda z podgromady anapsydów, zaliczanego do rodziny Millerettidae. Zamieszkiwał lasy deszczowe na terenie Południowej Afryki w okresie późnego permu, ok. 250 mln lat temu. Wyglądem przypominał obecne jaszczurki, jego masa wynosiła ok. 200 g, długość zaś 60 cm. Milleretta żywiła się owadami i innymi drobnymi kręgowcami. Mimo, iż była anapsydem, posiadała otwory z obu stron głowy.
 W
WMillerozaur (Millerosaurus) – rodzaj niewielkiego, wymarłego zauropsyda z podgromady anapsydów, zaliczanego do rodziny Millerettidae. Gad ten zamieszkiwał w późnym permie tereny Afryki Południowej, miał około 30 cm długości, i mimo że należał do anapsydów, otwory skroniowe, charakterystyczne dla synapsydów i diapsydów. Prawdopodobnie, podobnie jak dzisiejsze jaszczurki, millerozaur był owadożercą.
 W
WNotozaur (Nothosaurus) – rodzaj zauropteryga z rzędu notozaurów i rodziny Nothosauridae. Żył od wczesnego do późnego triasu na terenie współczesnej północnej Afryki, Azji, Europy.
 W
WPakasuchus – rodzaj niewielkiego krokodylomorfa z grupy Notosuchia żyjącego u schyłku wczesnej kredy na obecnych terenach Afryki. Został opisany w 2010 roku przez Patricka O'Connora i współpracowników w oparciu o niemal kompletny, bardzo dobrze zachowany szkielet. Odnaleziono również szczątki kilku innych osobników. Odkryto je w osadach ogniwa Namba w formacji Galula w basenie Rukwa Rift, w południowo-zachodniej Tanzanii, pomiędzy jeziorami Tanganika i Niasa. Dokładny wiek tej formacji pozostaje nieznany, jednak skamieniałości Pakasuchus pochodzą prawdopodobnie z warstw mających 99–115 mln lat.
 W
WProlacerta – rodzaj wczesnego archozauromorfa przypominającego sylwetką i rozmiarami współczesne jaszczurki. Odnaleziona we wczesnotriasowych osadach Afryki i Antarktydy. Dawniej uznawana za ogniwo pośrednie między prymitywnymi permskimi diapsydami, takimi jak Youngina, a jaszczurkami. Poparte to było brakiem łuku jarzmowego, ale późniejsze badania wykazały, że absencja tej struktury jest cechą obecną już u wspólnego przodka lepidozaurów i archozaurów. Sugerowany kinetyzm czaszki również został zanegowany. Prolacerta została później przeniesiona do grupy Archosauromorpha, do nowo utworzonego rzędu Prolacertiformes, który okazał się polifiletyczny. Obecnie umiejscawia się ją blisko kladu Archosauriformes obejmującego m.in. archozaury. Znana z licznego materiału, obejmujących zarówno pozostałości czaszki jak i szkieletu pozaczaszkowego. Jej młodszym synonimem jest Pricea longiceps Broom i Robinson 1948 znana również z zony Lystrosaurus. Również Kadimakara Bartholomai, 1979 z wczesnego triasu Australii może okazać się synonimem prolacerty.
 W
WProtozuch (Protosuchus) – rodzaj prymitywnego wczesnojurajskiego krokodylomorfa z grupy Protosuchia i rodziny Protosuchidae. Nazwa Protosuchus oznacza „pierwszy krokodyl” i odnosi się do faktu, że jest to jedno z najwcześniejszych zwierząt zaliczanych do Crocodylia sensu lato, odpowiadających Crocodyliformes.
 W
WRazanandrongobe – rodzaj drapieżnego archozaura żyjącego w środkowej jurze na terenie dzisiejszego Madagaskaru. Został opisany przez Maganuco, Dal Sasso i Pasiniego (2006) na podstawie niekompletnej kości szczękowej z trzema zębami oraz kilku izolowanych zębów, odkrytych w datowanych na baton osadach formacji Sakaraha w Prowincji Mahajanga. Autorzy opisu Razanandrongobe początkowo zaliczyli go jedynie do archozaurów, wskazując, że skamieniałości tego taksonu przypominają budową szczęki i zęby teropodów oraz niektórych krokodylomorfów. W 2017 roku opisano kolejne szczątki Razanandrongobe, w tym niemal kompletną kość przedszczękową z pięcioma zębami oraz fragment kości zębowej. W tej drugiej znajdował się ząb o długości ponad 15 cm – największy ze wszystkich zębów przypisywanych do Razanandrongobe. Ich budowa sugeruje, że Razanandrongobe żywił się twardym pokarmem, takim jak ścięgna i kości. Prawdopodobnie cechował się on stosunkowo szerokim i wysokim pyskiem – z przodu żuchwa była bezzębna. Rozmiar odnalezionych skamieniałości dowodzi, że był to jeden z największych drapieżnych archozaurów Gondwany.
 W
WRhabdognathus – rodzaj krokodylomorfa z rodziny Dyrosauridae żyjącego od późnej kredy do wczesnego paleocenu na obecnych terenach Afryki. Został opisany w 1930 roku przez Swintona w oparciu o kilka fragmentów żuchwy pochodzących z paleoceńskich osadów w Nigerii. Kolejne szczątki żuchwy przypisali do tego rodzaju Halstead i Middleton w 1976 roku, a w 1980 Éric Buffetaut opisał pierwsze skamieniałości czaszki. W 2002 roku Christopher Brochu i współpracownicy opisali puszkę mózgową, którą wstępnie przypisali do Rhabdognathus. Ze względu na fragmentaryczność holotypu zaliczenie innych szczątków czaszki do rodzaju Rhabdognathus nie mogło być całkowicie pewne. W 2007 roku Stéphane Jouve przeanalizował wszystkie znane skamieniałości należące do Rhabdognathus i opisany przez Swintona R. rarus uznał za nomen dubium, jednak utrzymał go jako gatunek typowy, stwierdzając, że – mimo iż niediagnostyczny na poziomie gatunkowym – pozwala na odróżnienie Rhabdognathus od innych rodzajów. Jouve nazwał także dwa nowe gatunki – R. aslerensis i R. keiniensis – a kolejny, R. compressus, został przeniesiony do rodzaju Congosaurus w oparciu o budowę zębodołów, sugerującą, że jest on bliżej spokrewniony z C. bequaerti.
 W
WSarkozuch (Sarcosuchus) – rodzaj wymarłego wczesnokredowego gada pokrewnego współczesnym krokodylom, ale niebędącego ich przodkiem.Opis: Stosunkowo wąskie i długie szczęki zawierały 132 masywne, stożkowate zęby. Pysk stanowi 3/4 długości całej czaszki. Szczęka trochę dłuższa od żuchwy. Wzdłuż grzbietu szereg dużych płytek kostnych o długości do 1 m. Wielkość: 11–12 m i waga około 8 ton, w tym czaszka 1,78 m. Był jednym z największych krokodylomorfów wszech czasów. Występowanie: Afryka. Ekologia rodzaju: Słodkowodny. Analiza kształtu zębów wskazuje, że gad używał ich nie do cięcia, a do bardzo mocnego trzymania ofiary i rozrywania dzięki np. potrząsaniu ciałem zdobyczy. Analiza szczęk i odcisków oraz przyczepów mięśniowych sugeruje nacisk szczęk o sile 80 tysięcy niutonów. Uważa się, że młodociane osobniki odżywiały się głównie rybami, ale starsze mogły polować także na duże żółwie i zwierzęta lądowe, w tym mniejsze dinozaury. Na podstawie analizy struktury kości szacuje się, że żyły przeciętnie 50–60 lat. Gatunki: S. imperator Broin & Taquet, 1966 S. hartii (Marsh, 1869) Muzea:
 W
WSimosuchus – rodzaj krokodylomorfa z grupy Notosuchia żyjącego w późnej kredzie na terenach Madagaskaru. Osiągał niewielkie rozmiary, nie dorastając do metra długości. Miał tępo zakończony, krótki pysk i wysoką, zaokrągloną czaszkę. Krępy tułów zakończony krótkim ogonem oraz kończyny pokrywał pancerz z osteoderm. Jego zęby przypominały zęby różnych roślinożernych kręgowców, w tym dinozaurów ptasiomiednicznych, co sugeruje, że odżywiał się głównie – jeśli nie wyłącznie – roślinami. Prowadził lądowy tryb życia, poruszał się na lekko ugiętych kończynach. Spekulowano, że mógł kopać nory, jednak brak na to jednoznacznych dowodów. Przypuszczalnie u Simosuchus występował dymorfizm płciowy – czaszki samic miałyby być krótsze i węższe, ale wyższe od czaszek samców.
 W
WSiroccopteryx – rodzaj pterozaura żyjącego na przełomie wczesnej i późnej kredy na terenach dzisiejszej Afryki. Gatunkiem typowym jest S. moroccensis, którego holotypem jest przednia część szczęki z zachowanymi zębami, odkryta niedaleko Beg'aa na południowy zachód od miasta Taouz w Maroku. Unwin (2001), Frey, Martill i Buchy (2003) oraz Ibrahim i współpracownicy (2010) uznali Siroccopteryx za młodszy synonim rodzaju Coloborhynchus, klasyfikując S. moroccensis jako odrębny gatunek w obrębie rodzaju Coloborhynchus; jednak zdaniem Rodrigues i Kellnera (2008) nie ma dowodów na to, że S. moroccensis reprezentuje ten sam rodzaj, co gatunek typowy rodzaju Coloborhynchus, C. clavirostris.
 W
WVoay – rodzaj wymarłego krokodyla z rodziny krokodyli właściwych (Crocodylidae). Mierząc 3–4 m długości i ważąc około 170 kg, był największym drapieżnikiem żyjącym w późnym plejstocenie i holocenie na Madagaskarze. Wyginął około 2000 lat temu, w przybliżeniu w tym samym czasie, kiedy na tej wyspie osiedlili się ludzie. Gatunkiem typowym jest Voay robustus, początkowo zaliczony do rodzaju Crocodylus. Nowsze analizy wskazują jednak na jego bliższe pokrewieństwo z krokodylem krótkopyskim niż z Crocodylus.